УДК 581.1 Подъем воды в ксилеме растений с трахеидальной проводимостью*
Как следует из целого ряда публикаций корневое давление, во-первых, не постоянно и зависит от концентрации минеральных веществ, влажности почвы, температуры и т.д. и, во-вторых, не превышает 0,15МПа для растений средней полосы при самых благоприятных условиях. Таким образом, корневое давление не является механизмом, обеспечивающим подъем воды в растениях. Полагая поверхностное натяжение в мениске капилляра ксилемы растений основной причиной подъема воды, необходимо учитывать особенности строения ксилемы хвойных и лиственных деревьев. В работе получено уравнение для расчета подъема воды в капилляре ксилемы растений с трахеидальной проводимостью. Уравнение выведено с учетом анатомического строения трахеид и окаймленных пор. Уравнение позволяет определить изменение гидравлических потерь при движении воды через одиночную окаймленную пору и через их совокупность с учетом ориентации последних.
Ключевые слова: ксилема, окаймленная пора. трахеида, ствол, растение, капилляр
Введение. Наличие вертикального восходящего потока воды в ксилеме растений объясняется действием силы поверхностного натяжения воды в менисках капилляров [1]. При объяснении подъема воды в капилляре радиуса r на высоту h традиционно рассматривают погруженный в воду стеклянный вертикальный капилляр и записывают условие равенства двух сил: силы, обусловленной весом столба воды в капилляре Р, направленной вертикально вниз, и направленной вверх силы поверхностного натяжения воды в мениске капилляра Fн. Уравнение равновесия этих сил для цилиндрического капилляра записывается в виде
Р = Fн ↔ πr2 hρg = 2π r σ cos α , (1)
где ρ , g , α , σ – соответственно плотность воды, ускорение свободного падения, угол смачивания и коэффициент поверхностного натяжения воды.
Из (1) следует известная формула Жюрена, позволяющая определить высоту подъема воды в вертикальном капилляре [2]
hж ![]() (2)
(2)
Применение формулы (2) для оценки высоты подъема воды в капиллярной системе древесных растений имеет тот недостаток, что при этом не учитывается сила сопротивления движению воды, обусловленная анатомическим строением капиллярной сети ксилемы а также сила, обусловленная осмотическим потенциалом корневой системы, приводящая к появлению действующего в сторону Fн корневого давления ркор, величина которого, согласно [3] , [4], для растений средней полосы при оптимальных условиях составляет 0,15 МПа. Капилляр ксилемы, по которому поднимается вода, например, у хвойных деревьев состоит из связанных посредством пор трахеид. В отличие от хвойных, ксилема лиственных деревьев состоит из сосудов, связанных между собой многочисленными порами, имеющимися на их поверхности. Существуют также ксилемы смешанного типа - трахеидально-сосудистые. Но и в этом случае связь между элементами таких ксилем осуществляется посредством пор. С учетом вышеизложенного соотношение (1) может быть записано следующим образом
Р + Fсп = Fн + Fкор ↔ πr2 hρg + Fсп = 2π r σ cos α + πr2 ркор , (3)
где
Fсп = ![]() i - суммарная
сила сопротивления движению воды,
обусловленная гидравлическими потерям напора при ее движении через сложную
капиллярную структуру, содержащую N пор, ркор - корневое
давление.
i - суммарная
сила сопротивления движению воды,
обусловленная гидравлическими потерям напора при ее движении через сложную
капиллярную структуру, содержащую N пор, ркор - корневое
давление.
До настоящего времени в литературных источниках отсутствуют сведения о гидравлических потерях напора как при движении воды через одиночную пору, так и через их совокупность в растении.
Среди большого разнообразия видов пор [5] наиболее хорошо и полно изучены и описаны геометрические размеры и строение округлых окаймленных пор с торусом, являющихся достаточно совершенным водопроводящим элементом. В структуре ксилемы хвойных деревьев трахеиды с окаймленными порами составляют 90 - 95 % от общего объема древесины. . Окаймленные поры расположены преимущественно на радиальных стенках у концов трахеид, которыми каждая трахеида вклинивается между соседними, образуя плотное соединение. Поры в стенках двух смежных трахеид располагаются одна против другой, образуя пару пор. На радиальной стенке трахеиды может находиться до 90 окаймленных пор, которые участвуют в проведении воды.
Цель работы. Расчет гидравлических потерь, возникающих при движении воды в ксилеме деревьев, как через одиночную окаймленную пору с торусом, так и через их совокупность и оценка влияния этих потерь на величину высоты подъема воды в капиллярах ксилемы.
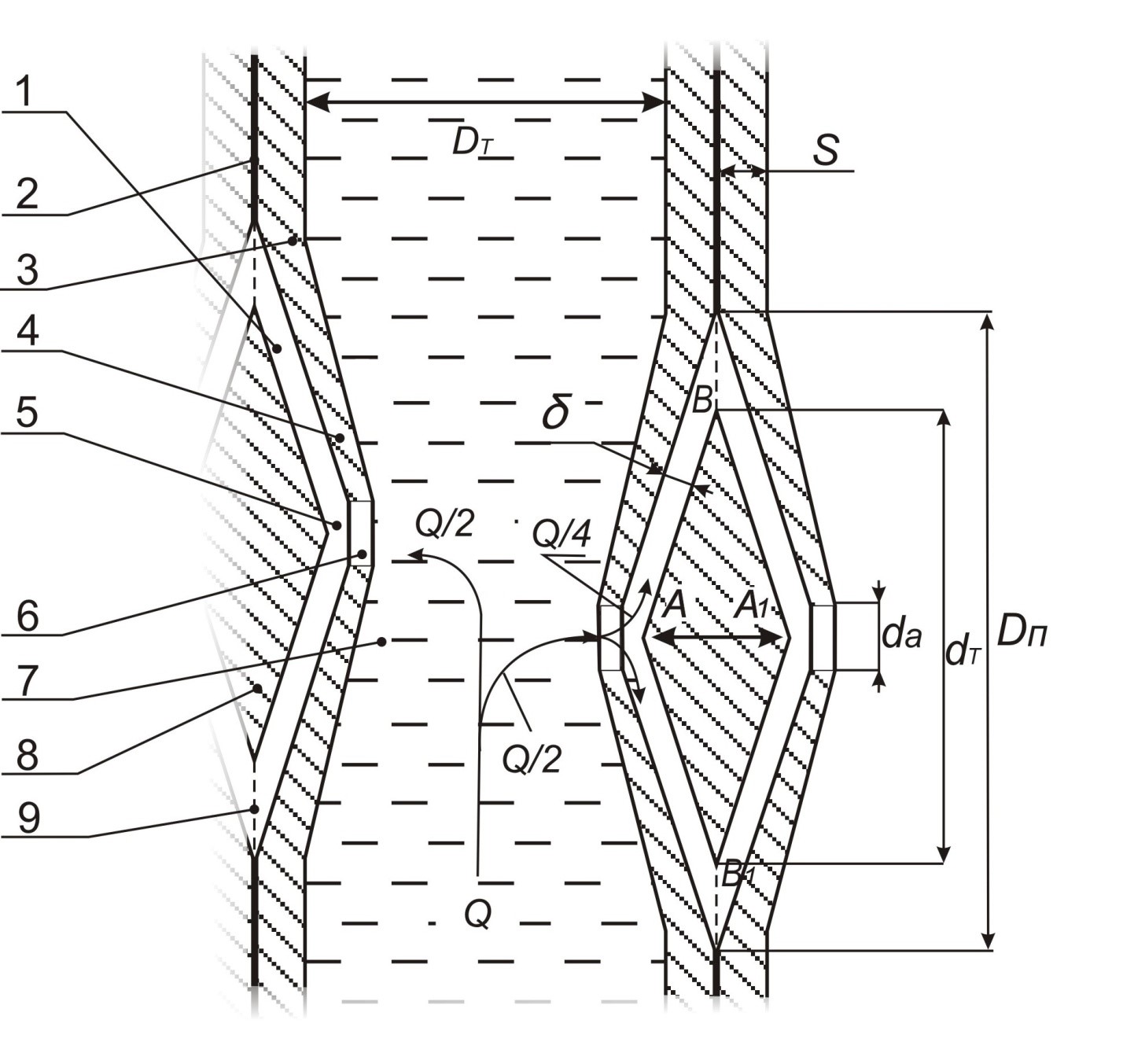
Основная часть. На рис.1 представлена модель участка трахеиды с внутренним диаметром Dт=2Rт, которая имеет на противоположных радиальных стенках по одной одинаковой окаймленной поре с округлым отверстием.
Окаймленные поры, образующиеся в стенках трахеид и сосудов, характеризуются тем, что полость поры суживается в направлении от замыкающей пленки к внутреннему отверстию поры. В окаймленной поре можно различить поровый канал 1, воронковидно расширяющийся по направлению к замыкающей пленке 2, и вторичную стенку 3, нависающую в виде валика (4) над расширенной частью канала и образующую камеру поры (5).
Канал сообщается внешним отверстием с камерой поры , а внутренним (апертура поры ) (6) - с полостью (7) самой трахеиды, что хорошо видно на фотографиях, выполненных через оптический и электронный микроскопы [6] (рис 2.75). Срединная часть замыкающей пленки окаймленной поры имеет чечевицеобразное утолщение, называемое торусом 8. Торус соединен с
замыкающей пленкой тонкими нитями (9), между которыми нет никакой перепонки. Поэтому часть замыкающей пленки поры, на которой подвешен торус не представляет собой сплошной перегородки и не создает препятствия при движении воды.
У ранних трахеид средний радиальный размер составляет для лиственных и хвойных пород соответственно 52· 10-6 и 40·10-6 м. а у поздних трахеид - 22· 10-6 и 20·10-6 м. Тангенциальный размер в обоих случаях примерно 30·10-6 м [7].
Диаметр окаймленных пор Dп колеблется от 8· 10-6 до 31·10-6 м, а диаметр входного и выходного отверстий в порах dа (апертура) от 4·10-6 до 8·10-6 м . Диаметр торуса dт всегда больше dа. Численные данные о соотношении диаметра торуса, диаметра поры и максимальной его толщине (АА1 на рис.1) в литературных источниках не приводятся. Толщина вторичной стенки поры s может достигать 10· 10-6 м.
Для определенности положим, что пора симметрична относительно вертикали и горизонтали и что зазор δ (поровый канал) между торусом и стенкой поры одинаков на всем пути движения воды, а максимальная толщина торуса равна 10· 10-6 м - толщине вторичной клеточной стенки. Тогда пути АВА1 = АВ1А1 = l , проходимые водой внутри поры в указанных выше пределах диапазона диаметров Dп , будут одинаковы и находиться в следующем
интервале: 5·10-6 ≤ l ≤ 30·10-6 м.
В силу одинаковости пор общий поток воды Q , передаваемый из трахеиды к смежным с ней трахеидам, делится пополам и в каждую пору поступает объем воды равный Q/2. В поровой камере вода обтекает торус восходящим и нисходящим потоками, каждый из которых составляет объем q= Q/4.
Величина Q зависит от скорости подъема воды в растении V, величина которой варьируется в широких пределах и зависит от вида растения, времени суток и сезона, в котором проводятся измерения. В [8] (стр. 386) приведены значения скоростей подъема воды в ксилеме растений, большинство значений которых лежит в пределах от 1 до 10 м/ч. В [9] (стр.112) приводятся данные, согласно которым в хвойных растениях вода передвигается с максимальной скоростью 1,2 м/ч, в лиственных значительно выше, а у дуба 27 – 40 м/ч.
Для определения величины потерь гидравлического напора воды при прохождении ее через окаймленную пору с торусом воспользуемся формулой Пуазейля, описывающей закономерность течения вязкой несжимаемой жидкости в тонкой цилиндрической капиллярной трубке [10]:
q = π d4 (p1 – p2) / (128 l μ) (4)
где:
q – секундный объемный расход воды , м3/с
p1 – p2 = Δр – перепад давления на входе и выходе из окаймленной поры, Па;
d – диаметр капилляра, м; μ – коэффициент динамичеcкой вязкости воды, Па·с ;
l – длина капилляра, м.

Рис.1. Модель участка трахеиды с окаймленной порой
на противоположных радиальных стенках.
Из (4) следует:
p1 – p2 = Δр = 128 l μ q / (π d4) (5)
Величину d в (5) выберем из следующих соображений. Поступающая через апертуру в пору вода движется на участке ВАВ1 вдоль конического зазора. При этом по мере ее продвижения количество воды, приходящееся на единицу площади поперечного сечения зазора, уменьшается, а, следовательно, уменьшается скорость ее движения и гидравлические потери. Но на участке ВА1В1 коническое расширение сменяется противоположным уменьшением площади сечения зазора, приводящим к увеличению скорости движения воды и величины гидравлического сопротивления. В результате в силу симметрии гидравлические потери будут определяться только величиной пути, по которому движется в зазоре вода. Поэтому движение воды в зазоре можно рассматривать как происходящее в капилляре длиной l при d = δ. Исходя из возможных соотношений Dп и dа зададим диапазон изменения δ: 2 ·10-6 ≤ δ ≤ 8·10-6м. Примем также диапазон изменения V следующим: 0,28·10-3 ≤ V ≤ 2,2·10-3 м/с.
В таблице 1 приведены величины объема воды q
= Q/4 (Q = π![]() V/4), проходящего
за секунду через зазор δ по пути АВА1
между поверхностью торуса и внутренней поверхностью вторичной стенки окаймленной поры для трахеид диаметрами Dт равными 20·10-6 , 30·10-6 и 40·10-6 м и при разном количестве пор N на смежных
стенках трахеид, через которые проходит вода. При этом в каждом случае скорость подъема
воды в ксилеме V принимает значения 1, 4 и 8 м/ч.
V/4), проходящего
за секунду через зазор δ по пути АВА1
между поверхностью торуса и внутренней поверхностью вторичной стенки окаймленной поры для трахеид диаметрами Dт равными 20·10-6 , 30·10-6 и 40·10-6 м и при разном количестве пор N на смежных
стенках трахеид, через которые проходит вода. При этом в каждом случае скорость подъема
воды в ксилеме V принимает значения 1, 4 и 8 м/ч.
Можно видеть, что секундный объем воды, протекающий через одиночную пору, уменьшается прямо пропорционально N и увеличивается прямо пропорционально V.
Таблица 1
|
Dт ·106, м |
V · 103,м/с |
q 1013, м3/с | |||
|
N | |||||
|
1 |
10 |
25 |
50 | ||
|
20 |
0,28 |
0,22 |
0,022 |
0,0088 |
0,0044 |
|
1,10 |
0,87 |
0,087 |
0,0348 |
0,0174 | |
|
2,20 |
1,74 |
0,174 |
0,0697 |
0,0350 | |
|
30 |
0,28 |
0,50 |
0,050 |
0,0198 |
0,0100 |
|
1,10 |
1,96 |
0,196 |
0,0780 |
0,0390 | |
|
2,20 |
3,90 |
0,390 |
0,1550 |
0,0770 | |
|
40 |
0,28 |
0,87 |
0,087 |
0,0350 |
0,0178 |
|
1,10 |
3,50 |
0,350 |
0,1390 |
0,0700 | |
|
2,20 |
7,00 |
0,700 |
0,2790 |
0,1400 | |
На рис 2 приведены рассчитанные по формуле (5) величины потерь давления Δрмин при прохождении водой окаймленной поры при минимальных значениях величин Vмин =0,28·10-3м/с, Nмин,=1, lмин= 5·10-6м и δмин=2·10-6м, принятых в рассматриваемой нами модели, для трахеид диаметрами 20·10-6 ,30·10-6 и 40·10-6 м. Из рисунка следует, что при прохождении воды через одиночную пору с δмин= 2·10-6 м величина Δрмин возрастает при увеличении диаметра трахеиды.
На основании (5) могут быть определены величины Δр для других значений указанных выше величин по следующим соотношениям.
Увеличение l приводит к увеличению потерь давления, то есть
![]() =
= ![]() (6).
(6).
Увеличение количества окаймленных пор, участвующих в проведении воды, приводит к уменьшению потерь давления, а именно
![]() =
= ![]() (7).
(7).
Δрмин
Па

Dт·10-6 м
Рис.2. Зависимость Δрмин от диаметра трахеиды Dт.
Увеличение скорости продвижения воды в
трахеидах ксилемы увеличивает потери давления , что описывается соотношением
![]() =
= ![]() (8)
(8)
Увеличение величины зазора δ приводит к изменению потерь давления описываемого соотношением
![]() =
=  (9).
(9).
С учетом данных таблицы 1 и рис 2, по соотношениям (6) – (9) проведены расчеты величины диапазона изменения Δр при изменении l в пределах 5·10-6 ≤ l ≤ 30·10-6 м , а величин V, δ в пределах: 0,28·10-3 ≤ V ≤ 2,2·10-3 м/с , и 2 ·10-6 ≤ δ ≤ 8·10-6м. Результаты расчета для трахеид диаметрами Dт равными 20·10-6м и 40·10-6м при движения воды через одиночную пору (N=1) и через 50 (N=50) одновременно проводящих воду пор на стенке трахеиды, представлены в таблице 2.
Как видно из таблицы 2 при N=1 величина Δр растет при увеличении l, V и Dт. Увеличение δ при прочих равных условиях приводит к уменьшению Δр. Абсолютная величина Δр в пределах рассматриваемого изменения параметров варьирует в широких пределах от 1,1 до 53200 Па. При этом потери
уменьшаются с ростом N , а максимальные их значения имеют место при N=1.
Таблица 2
|
N |
Dт ·106, м |
V, ·103 м/с |
δ·106,м
|
Интервал изменения Δр, Па при 5·10-6 ≤ l ≤ 30·10-6 м |
|
1 |
20
|
0,28 |
2 |
281 ÷ 1690 |
|
8 |
1,1 ÷ 6,6 | |||
|
2,2 |
2 |
2220 ÷ 13340 | ||
|
8 |
8,7 ÷ 52 | |||
|
40 |
0,28 |
2 |
2200 ÷ 13200 | |
|
8 |
4,4 ÷ 26,3 | |||
|
2,2 |
2 |
8940 ÷ 53200 | ||
|
8 |
35 ÷ 210 | |||
|
50 |
20 |
0,28 |
2 |
5,6 ÷ 34 |
|
8 |
0,02 ÷ 1,3 | |||
|
2,2 |
2 |
45 ÷ 268 | ||
|
8 |
0,17 ÷ 1,05 | |||
|
40 |
0,28 |
2 |
22 ÷ 135 | |
|
8 |
0,09 ÷ 0,52 | |||
|
2,2·10-3 |
2 |
179 ÷ 1080 | ||
|
8 |
0,7 ÷ 4,2 |
При увеличении количества пор на стенке трахеиды до 50 величина Δр значительно уменьшается и варьируется в пределах 0,02 до 1080 Па.
Как было отмечено выше, движение воды в коническом зазоре окаймленной поры в силу симметрии физических процессов можно рассматривать, как движение воды в капилляре длиной l и диаметром δ. Действительно, если принять за положительное направление действующих сил в сторону силы поверхностного натяжения, для участков АВ, ВА1, АВ1, В1А1 можно записать следующее уравнение для равнодействующей силы Fрп , обусловленной распределением веса воды в зазоре (Рис.3а)
Fрп= - ΔР sin (90 - ![]() ) + ΔР sin (90 -
) + ΔР sin (90 - ![]() ) + ΔР sin (90 -
) + ΔР sin (90 - ![]() ) - ΔР sin (90 -
) - ΔР sin (90 - ![]() ) = 0 (10)
) = 0 (10)
То есть на выходе из поры Fрп равно нулю и в этом смысле пора как бы «нейтральна».
Ситуация изменяется при отсутствии симметрии. Если повернуть оси по часовой стрелке на угол γ/2, то стороны ВА1 и АВ1 примут вертикальное положение (Рис.3б) и в сторону силы поверхностного натяжения будет действовать вес двух столбиков воды равный 2 ΔР. В то же время стороны АВ и В1А1 окажутся расположенными под углом γ к горизонтали. Тогда для
равнодействующей силы на выходе воды из поры можно записать соотношение
Fрп= - ΔР sin (90 - γ ) + ΔР + ΔР - ΔР sin (90 - γ ) = 2 ΔР (1 - sin (90 - γ )) ≠ 0 (11)
Трахеида не является вертикальным цилиндром, а клиновидно сужается радиальными стенками к концам, одной третью поверхности соприкасаясь с наклоненной под углом к вертикали окаймленной поры (б)

Рис.3 Распределение сил в зазоре вертикально ориентированной (а) и
клиновидными поверхностями концов трахеид следующих уровней [7] и поэтому вертикальная ось симметрии поры может быть наклонена. Угол γ клина на конце трахеиды лежит в пределах 17 – 22 градусов. Величина Fрп в одиночной поре в зависимости от γ может принимать значения в пределах
Fрп =
(0,015 ÷ 0,035) ΔР =(0,015÷0,035)πδ2ρg
l /8
![]() (0,002 ÷ 0,0044) πδ2 ρg
l. (12)
(0,002 ÷ 0,0044) πδ2 ρg
l. (12)
Тогда при минимальном и максимальном количестве трахеид на одном погонном метре величина Fрп соответственно равна (0,33 ÷ 9 ) πδ2 ρg l и (8 ÷ 19 ) πδ2 ρg l.
С учетом вышеизложенного уравнение (3) может быть записано в виде
Р + Fсп - Fрп = Fн + Fкор ↔ πr2 hρg + Fсп -Fрп = 2π r σ cos α + πr2 ркор (13)
Подставляя в (13) аналитические соотношения для величин Fсп = π ![]() Δр и Fрп из (11) и
полагая, что в стволах деревьев хвойных пород длина трахеид Δh находится в
диапазоне от 2,5·10-3м до 5·10-3м [7] получим
Δр и Fрп из (11) и
полагая, что в стволах деревьев хвойных пород длина трахеид Δh находится в
диапазоне от 2,5·10-3м до 5·10-3м [7] получим
π ![]() hρg
+ 32 π
hρg
+ 32 π ![]() l
μ V / N δ4 -
l
μ V / N δ4 - ![]() πδ2 ρg
l(1 - sin (90 - γ
) =
πδ2 ρg
l(1 - sin (90 - γ
) =
= 2π RТ σ cos α + π![]() ркор (14) Сокращая (14) на π
ркор (14) Сокращая (14) на π ![]() ρg и учитывая (2)
получим
ρg и учитывая (2)
получим
![]() = hж
= hж ![]() (15)
(15)
откуда
h = (hж![]() )
/
)
/ ![]() (16)
(16)
Полученное уравнение (16) позволяет определить высоту подъема воды в капилляре ксилемы, состоящей из вертикальной цепочки трахеид с учетом анатомического строения окаймленных пор.
Уравнения (13) и (14) позволяют выяснить роль и степень участия корневого давления в подъеме воды в ксилеме. Для этого составим отношение сил
Fсп/Fкор
= 32 π ![]() l
μ V / N δ4 π
l
μ V / N δ4 π![]() ркор = 32
ркор = 32 ![]() h
l
μ V / Δh N δ4 ркор (17)
h
l
μ V / Δh N δ4 ркор (17)
Если Fсп/Fкор = 1 , то вода под действием силы Fн поднимается на высоту hж , так как корневое давление компенсирует гидравлические потери при движении воды через поры.
При Fсп /Fкор < 1 корневое давление способствует подъему воды и вода поднимается на высоту большую hж , а при Fсп /Fкор > 1 часть Fн расходуется на преодоление Fсп и вода не может достичь hж. Записав (17) в виде
![]() ≤ ркор
≤ ркор ![]()
![]() (18) и, полагая
ркор = 0,15 МПа, можно определить, при каких параметрах справедливо (18). Так при RТ =10 ·10-6
м,
l = 5·10-6 м, μ= 1,002 ·10-3 Па·с,
V= 0,28·10-3м/с, δ = 2·10-6м, Δh= 2,5·10-3м
(18) и, полагая
ркор = 0,15 МПа, можно определить, при каких параметрах справедливо (18). Так при RТ =10 ·10-6
м,
l = 5·10-6 м, μ= 1,002 ·10-3 Па·с,
V= 0,28·10-3м/с, δ = 2·10-6м, Δh= 2,5·10-3м ![]() ≤ 1,34 N. Тогда при N=1 и при N=50 высота, ксилемы,
на которой происходит компенсация корневым давлением Fсп составляет
соответственно 1,34 и 66,9 м. При δ
= 8·10-6м и прочих равных
условиях
≤ 1,34 N. Тогда при N=1 и при N=50 высота, ксилемы,
на которой происходит компенсация корневым давлением Fсп составляет
соответственно 1,34 и 66,9 м. При δ
= 8·10-6м и прочих равных
условиях ![]() ≤ 341 N. При RТ =20 ·10-6 м, l = 30·10-6
м, V=
2,2·10-3м/с, δ = 2·10-6м, Δh= 5·10-3м
≤ 341 N. При RТ =20 ·10-6 м, l = 30·10-6
м, V=
2,2·10-3м/с, δ = 2·10-6м, Δh= 5·10-3м ![]() ≤ 0,014N , а при δ =
8·10-6м
≤ 0,014N , а при δ =
8·10-6м
![]() ≤ 3,63N. Задавая N можно определить h.
≤ 3,63N. Задавая N можно определить h.
Большая вариативность переменных в (18) не позволяет перебором параметров наглядно представить результаты возможных вариантов их сочетания. Решить это можно табулированием результатов расчета. Отметим следующее. Каждая трахеида одной своей третью вклинивается радиальными стенками между радиальными стенками двух трахеид следующего уровня, где и расположена основная масса окаймленных пор [7]. При этом на одном погонном метре может быть размещено от 260 до 530 трахеид (в среднем 395). Полагая, что все трахеиды по высоте ксилемы имеют одинаковую длину и используя данные таблицы 2, можно определить диапазон потерь давления на один погонный метр трахеидального капилляра ксилемы. Результаты такого расчета представлены в таблице 3.
Как было отмечено выше, максимальная величина ркор, , не превосходит 0,15 МПа. Задавая условные высоты, например, 100, 50 и 10 метров м, из данных таблицы 3 можно определить интервалы изменения величин входящих в (18), в которых Δр превышает ркор в капиллярах разной высоты. При этом жирным шрифтом, курсивом с подчеркиванием и скобками выделены те значения Δр в рассматриваемом интервале изменения l, в которых Δр не превышает ркор при высотах капилляров соответственно 100 , 50 и 10 метров. В остальных случаях
Таблица 3.
|
N |
Dт·106 м |
V ·103, м/с |
δ, ·106 м
|
Интервал изменения Δр,МПа при 5·10-6 ≤ l ≤ 30·10-6 м и количестве трахеид на 1м | ||
|
260 (минимум) |
395 (среднее) |
530 (максимум) | ||||
|
1 |
20
|
0,28 |
2 |
0,073- 0,439 |
0,111 – 0,667 |
0,149 – 0,895 |
|
8 |
0,000286 – 0,0018 |
0,00043 – 0,0026 |
0,0006 –(0,0035) | |||
|
2,2 |
2 |
0,572 – 3,468 |
0,087 – 5,27 |
1,77 – 7,07 | ||
|
8 |
0,0022 – (0,0045) |
(0,0034)– 0,021 |
(0,0049) – 0,029 | |||
|
40
|
0,28 |
2 |
0,572 – 3,468 |
0,87 – 5,21 |
1,78 – 7,07 | |
|
8 |
0,0011 – (0,0068) |
0,0018 – (0,011) |
0,0023 –(0,0138) | |||
|
2,2 |
2 |
2,324 – 13,83 |
3,53 – 21,0 |
4,74 – 28,19 | ||
|
8 |
(0,0091) – 0,055 |
(0,014) – 0,083 |
0,0186 – 0,111 | |||
|
50 |
20 |
0,28 |
2 |
0,0014 – (0,0088) |
0,0022 – (0,013) |
0,00297- 0,018 |
|
8 |
0,000005 – 0,0003 |
0,00008 – 0,0005 |
0,00001 – 0,0007 | |||
|
2,2 |
2 |
(0,0117) – 0,069 |
0,018 – 0,106 |
0,024 – 0,142 | ||
|
8 |
0,00004 – 0,00027 |
0,00007 – 0,0004 |
0,0009 – 0,0006 | |||
|
40 |
0,28 |
2 |
(0,0057) – 0,035 |
(0,0087) – 0,053 |
0,0012 – 0,072 | |
|
8 |
0,000023 – 0,00014 |
0,000036 – 0,0002 |
0,00005 – 0,0003 | |||
|
2,2 |
2 |
0,046 – 0,28 |
0,071 – 0,427 |
0,0948 – 0,572 | ||
|
8 |
0,000182 – 0,001 |
0,00028 – 0,0017 |
0,00037 – 0,0022 | |||
(шрифт без выделений) величина Δр либо значительно превосходит ркор, либо равенство возможно при меньших высотах капилляра.
Анализ данных таблицы 3 показывает следующее. При малых δ и N=1 величина Δр растет с увеличением l, V и Dт уменьшаясь при увеличении длины трахеиды и уже на одном погоном метре капилляра находится в пределах 0,073 ÷ 28,19 МПа, что значительно превышает ркор. По мере увеличения δ при прочих равных условиях величина Δр уменьшается и при δ = 8·10-6 м находится в диапазоне 0,000286 ÷ 0,111 МПа, увеличиваясь прямопропорционально l , также уменьшаясь при увеличении длины трахеиды Δh .
При увеличении N поток воды через каждую окаймленную пору уменьшается, что приводит к снижению Δр. Так при N=50 и δ = 2·10-6 м Δр на одном погонном метре капилляра находится в пределах 0,0014 ÷ 0,572 МПА и для ряда значений l, V и Dт и длины трахеиды Δр ≤ ркор. При δ = 8·10-6 м Δр < ркор и находится в пределах 0,000005 ÷ 0,0022 МПа.
Малая величина δ характерна для мелких пор с Dп порядка 8· 10-6 м, где с учетом размера торуса , который не может быть меньше dа , зазор δ между ним и стенкой поры не может быть больше 2·10-6 м и, следовательно, Δр велико и ксилема с таким размером пор не может поднимать воду на большую высоту. Для крупной поры с Dп = 30· 10-6 м и dа =8· 10-6 м величина δ может достигать 8· 10-6 м .При этом Δр не велико и ксилема с такими порами на стенках трахеид может поднимать воду на большую высоту. Этот результат хорошо согласуется с [7], где сказано, что на радиальных стенках, особенно у концов ранних трахеид, имеется от 70 до 90 крупных окаймленных пор с округлыми отверстиями.
Следует заметить, что корневое давление не является постоянной величиной, а зависит от условий жизнедеятельности корня и таких факторов, как наличие и концентрация в почвенном растворе и клетках корня минеральных веществ, воды, температуры, недостатке кислорода, времени суток, сезона и др. и может быть значительно ниже 0,15 МПа. Поэтому корневое давление, частично или полностью компенсирует Δр, позволяя силам поверхностного натяжения в капиллярах ксилемы поднять воду на максимально возможную высоту. И если корневое давление падает, например, при недостатке влаги в почве, то силам поверхностного натяжения приходится преодолевать силы сопротивления, и они не могут поднять воду на прежнюю высоту, а растение начинает увядать с верхних уровней, где дефицит воды будет больше.
Выводы.
1.Получено уравнение, позволяющее определить высоту подъема воды в капилляре ксилемы растений с трахеидальной проводимостью с учетом анатомического строения трахеид и окаймленных пор с учетом ориентации последних. Показано, что движение воды через окаймленную пору можно рассматривать, как течение воды через капилляр длиной равной пути, проходимого водой при обтекании торуса и диаметром равным величине зазора между поверхностью торуса и внутренней стенкой трахеиды.
2. Определены пределы изменения гидравлических потерь при движении воды, как через одиночную окаймленную пору, так и через их совокупность в зависимости от зазора и длины проходимого водой пути в поре. Показано, что сопротивление одиночной окаймленной поры велико и при скоростях подъема воды в ксилеме 1 – 8 м/ час находится в пределах 0,073 – 28 МПа. При увеличении количества пор на стенке трахеиды гидравлические потери уменьшаются и составляют для тех же скоростей движения воды 0,000005 – 0,057 МПа.
3. Увеличение диаметра полости трахеиды и диаметра окаймленной поры и торуса при прочих равных условиях приводит к росту гидравлических потерь.
4. Увеличение как длины трахеиды, так и зазора между поверхностью торуса и поверхностью внутренней стенки трахеиды при прочих равных условиях приводит к снижению гидравлических потерь. Последнее обусловлено снижением скорости потока в зазоре.
5. При наклонном положении поры на стенке трахеиды необходимо учитывать направленную в сторону подъема воды дополнительную силу, возникающую из-за перераспределения веса воды, действующего на поверхность торуса и стенки поры.
6. Основной силой, осуществляющей подъем воды в ксилеме растений, является сила поверхностного натяжения в менисках капилляров.
7. Полученные результаты позволяют предположить, что при длительном дефиците воды или высокого содержания солей в почвенном растворе корневое давление может быть мало, близко к нулю и даже отрицательно и силам поверхностного натяжения приходится преодолевать сопротивление движению воды окаймленными порами. В результате вода в ксилеме не может подниматься на прежнюю высоту и начинается увядание, которое должно происходить, начиная от наиболее удаленных от комля концов наиболее длинных ветвей верхних ярусов растения.
Библиографический список.
1. Кипнис И.А. Механизм продвижения воды в капиллярах ксилемы растений. /И.А.Кипнис, Ю.М.Вернигоров. Вестник Дон. гос. техн. ун-та, 2014.-Т.14, №3 (78), - С.78-88.
2. Шеин Е.В. Курс физики почв./ Е.В.Шеин. – М.: МГУ, 2005.- 432 с.
3. Малиновский В.И. Физиология растений. / В.И. Малиновский. – Владивосток, изд. ДВГУ, 2004. 109 с.
4. Кузнецов Вл.В., Дмитриева Г.А. Физиология растений. /Вл.В. Кузнецов, Г.А.Дмитриева. Изд. 2-е, перераб. и доп. – М.: Высшая шк., 2006. -742 с.
5. Курсанов Л.И. Ботаника (в двух томах). Том 1. Анатомия и морфология. Для педагогических институтов и университетов. /Курсанов Л.И., Комарницкий Н.А., Мейер К.И., Раздорский В.Ф., Уранов А.А. Изд. 5-е., переработ. М.: Просвещение, 1966. 423с.
6. Ботаника. Учебник для вузов: в 4 т./П.Зитте, Э.В. Вайлер, Й.В. Кадерайт, А. Брезински, К. Кёрнер; на основе учебника Э. Страсбургера [ и др.]; пер. с немецкого – М. Издательский центр «Академия», 2008. –367 с. Т.1. Клеточная биология. Анатомия. Морфология./ под ред. А.К. Тимонина и В.В. Чуба.
7. Уголев Б.Н. Древесиноведение и лесное товароведение: учебник для студ. сред. проф. образования / Б.Н. Уголев. – 3-е изд., стер.- М. : Издательский центр «Академия», 2010. – 272 с.
8. Крамер Пол Д. Физиология древесных растений./ Пол Д. Крамер, Теодор Т. Козловски Пер. с англ. – М.: «Лесная промышленность», 1983 – 464 с.
9. Якушкина Н.И, Бахтенко Е.Ю. Физиология растений. /Н.И. Якушкина, Е.Ю. Бахтенко.- М.: Гуманитар. Изд. Центр ВЛАДОС, 2005. – 463 с.
10. Яворский Б.М. Справочник по физике / Б.М.Яворский, А.А. Детлаф.- Изд. «Наука», 1968. - 940 с.
References.
1. Kipnis I.A., Vernigorov Yu.M Mechanism of promotion of water in the xylem capillaries plants. Bulletin of the Don State Technical University, 2014.-Т.14, №3 (78), pp.78-88.
2. Shein E.V. Rates of soil physics. / E.V.Shein. - Moscow: Moscow State University, 2005. – 432 р.
3.Malinovsky V.I. Plant Physiol. / V.I. Malinovsky. - Vladivostok, ed. FESU 2004. - . 109 p.
4. Kuznetsov Vl.V., Dmitrieva G.A. Plant Physiol. / Vl. V. Kuznetsov, G.A. Dmitrieva. Ed. 2nd, revised. and add. - M.: Higher wk., 2006. -742 p.
5. Kursanov L.I. Botany (in two volumes). Volume 1. Anatomy and morphology. For pedagogical institutes and universities. / L.I. Kursanov, N.A. Кomarnicki, K.I. Meyer, V.F. Razdorskaya, A.A. Urunov. Ed. 5-that is, the overtime." M: Education, 1966 - 423 р.
6. Botany. Textbook for high schools: in 4 volumes :/ P.Zitte, E.V. Weiler, J.W. Kaderayt, A. Brezinský, C. Koerner, based on the textbook E. Strasburger [et al.], Trans. with German. Volume 1 Cell biology. Anatomy. Morphology./ edited by A.K. Timonina CENTURIES and Chub - Moscow: Publishing Center "The Academy", 2008 – 367 р.
7. Ugolev B.N. Wood Science and Forestry Commodity: a textbook for stud. Environments. Prof. Education / B.N. Ugolev. - 3rd ed., Sr. - M.: Publishing Center "Academy", 2010. – 272 p.
8. Kramer Paul D.. Physiology of woody plants./ Paul D. Kramer , Theodore T. Kozlowski. Trans. from English. - M.: "Forestry", 1983 - 464 p.
9. Yakushkina N.I. Plant physiology: a textbook for students / N.I. Yakushkina,, E. Bahtenko. - M. Humanitarian. Ed. Center VLADOS 2005. - 463 p.
10. Jaworski B.M. Рhysics Handbook / M. Jaworski, A.A. Detlef.- Ed. "Science", 1968. - 940 р.
THE RISE OF WATER IN THE XYLEM OF PLANTS WITH TRACHEIDAL
CONDUCTIVITY
I.A.Kipnis, Yu.M. Vernigorov
As follows from a number of publications, root pressure, firstly, is not constant and depends on the concentration of mineral substances, soil moisture, temperature, etc. and, secondly, does not exceed 0,15 MPa for plants of the middle band under the most favorable conditions. Thus the root pressure is not a mechanism for ensuring that the ascent of water in plants. Assuming the surface tension of the meniscus of the capillary xylem of plants the main reason for the rise of water, it is necessary to take into account the structural features of the xylem of conifers and deciduous trees. In the present paper we have derived an equation for calculating the rise of water in capillary xylem of plants with tracheidal conductivity. The equation is derived taking into account the anatomical structure of tracheids and bordered pores. The equation allows us to determine the change in hydraulic losses in the movement of water through a single-lined pore and through their combination, taking into account the orientation of the latter.
Keywords: xylem, bordered pore. tracheid, trunk, plant, capillary.
Авторы
И. А. Кипнис, Ю. М. Вернигоров
*Работа выполнена в рамках инициативной НИР.

